Difference between revisions of "Team:Peking/Model"
| Line 289: | Line 289: | ||
<div class="mdl-card__supporting-text"> | <div class="mdl-card__supporting-text"> | ||
| + | <h1>Control Unit</h1> | ||
| + | <h2>Intention of the Model</h2> | ||
| + | |||
| + | In this model, we aim to describe the sequence | ||
| + | modification process of the control unit sequence with recombination sites. Different | ||
| + | recombinases are expressed in series to alter the direction of terminators between | ||
| + | corresponding recombination sites on the control unit circuit. The combination of | ||
| + | unidirectional terminators in different directions (reverse or foward) determines the state | ||
| + | of RNA polymerase flux, thus controlling expression of the downstream gene.<br><br> | ||
| + | |||
| + | Though we have done simulation of recombinase dynamics in the previous section, it is | ||
| + | necessary to consider two key concepts of biological processes:<br><br> | ||
| + | 1. population behavior | ||
| + | 2. stochastic process | ||
| + | |||
| + | <br><br>The reason for modeling cell population behavior is that deployment of synthetic | ||
| + | bacterial | ||
| + | devices would certainly involve populations of cells. Furthermore, we demonstrate our design | ||
| + | through continuous imaging of entire cell cultures. This is a theory-experiment interaction | ||
| + | procedure. The reason for stochastic modeling is self-evident. Cellular biological | ||
| + | processes, especially gene expression, are innately noisy and stochastic.[^Footnote1] | ||
| + | Previous work has demonstrated the effectiveness of the integration of population-level | ||
| + | dynamics and stochastic cellular responses to inputs.[^Footnote2]It is important to explain | ||
| + | how stochastic single-cell responses affect overall population-level distributions and | ||
| + | behavior. | ||
| + | <br><br>This section describes a Markov model of recombinase-based DNA flipping and a Gillespie | ||
| + | stochastic simulation method[^Footnote3] to observe theoretical sequence state evolution. We | ||
| + | will focus on our demonstration design, which can be generalized to other designs.<br><br> | ||
| + | |||
| + | <h2 id="biological-basis">Biological Basis</h2> | ||
| + | The control units are implemented experimentally by transforming a plasmid into an E. coli. | ||
| + | This plasmid has similar structure as the reporter plasmid in the previous section. However, | ||
| + | in this situation the sequence fragments being inverted are terminators. Using | ||
| + | experimentally screened uni-directional terminators, the transcription or polymerase | ||
| + | flux[^Footnote4] can be precisely controlled. The figure below illustrates the result of the | ||
| + | design software. (超链接到software) | ||
| + | <img src="./1509366096800.png" alt="Alt text"/> <img | ||
| + | src="./1509366027555.png" alt="Alt text"/> | ||
</div> | </div> | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
<div class="mdl-card__menu"> | <div class="mdl-card__menu"> | ||
<button class="mdl-button mdl-button--icon mdl-js-button mdl-js-ripple-effect"> | <button class="mdl-button mdl-button--icon mdl-js-button mdl-js-ripple-effect"> | ||
| Line 308: | Line 342: | ||
<div class="mdl-card__supporting-text"> | <div class="mdl-card__supporting-text"> | ||
| − | <h1>Clock</h1> | + | <h1>Clock</h1><br> |
| − | < | + | |
| + | <section class="docs-toc docs-text-styling" style="margin-left: 0px"> | ||
| + | <nav class="section-content" style="margin-left: 0px"> | ||
| + | <ul style="margin-left: 0px;"> | ||
| + | |||
| + | <li><a href="#p1">Biological Basis and Assumptions</a> | ||
| + | </li> | ||
| + | |||
| + | <li><a href="#p2">Equations and Parameters</a> | ||
| + | </li> | ||
| + | |||
| + | |||
| + | |||
| + | <li><a href="#p3">Simulation Results</a> | ||
| + | </li> | ||
| + | |||
| + | </ul> | ||
| + | </nav> | ||
| + | </section> | ||
| + | |||
| + | <H2>Intention of the Model</H2> | ||
We attempts to develop a framework of biological sequential circuits that are programmable. | We attempts to develop a framework of biological sequential circuits that are programmable. | ||
The envisioned circuit is capable of both storing states in DNA and automatically running a | The envisioned circuit is capable of both storing states in DNA and automatically running a | ||
| Line 323: | Line 377: | ||
possibility of repressilator-driven bio-flip-flop. | possibility of repressilator-driven bio-flip-flop. | ||
| + | </div> | ||
| + | </div> | ||
| + | <div id = "cl1" class="demo-card-wide mdl-card mdl-shadow--2dp"> | ||
| + | |||
| + | <div class="mdl-card__supporting-text"> | ||
| − | + | <h2>Biological Basis and Assumptions</h2> | |
The | The | ||
repressilator is a synthetic genetic regulatory network consisting of a ring-oscillator with | repressilator is a synthetic genetic regulatory network consisting of a ring-oscillator with | ||
| Line 333: | Line 392: | ||
feedback loop, such that each gene represses the next gene in the loop, and is repressed by | feedback loop, such that each gene represses the next gene in the loop, and is repressed by | ||
the previous gene.<br><br> | the previous gene.<br><br> | ||
| − | <div align="center"><img src="https://static.igem.org/mediawiki/2017/8/80/Peking_modeling_fig_1.png" height="130px"/><br> | + | <div align="center"><img src="https://static.igem.org/mediawiki/2017/8/80/Peking_modeling_fig_1.png" |
| − | + | height="130px"/><br> | |
| + | <strong>Fig.1. The genetic structure of the repressilator network</strong></div> | ||
<br><br> | <br><br> | ||
We decided to implement our repressilator-triggered bio-flip-flop with two repressor proteins | We decided to implement our repressilator-triggered bio-flip-flop with two repressor proteins | ||
| Line 340: | Line 400: | ||
illustrated as follows:<br><br> | illustrated as follows:<br><br> | ||
| − | <div align="center"><img src="https://static.igem.org/mediawiki/2017/4/4a/Peking_modeling_fig_2.png" height="300px"/><br> | + | <div align="center"><img src="https://static.igem.org/mediawiki/2017/4/4a/Peking_modeling_fig_2.png" |
| − | + | height="300px"/><br> | |
| − | + | <strong>Fig.2. Schematic | |
| + | drawing of the repressilator-driven bio-flip-flop </strong></div> | ||
| + | <br><br> | ||
The upper promoter (X) is <span | The upper promoter (X) is <span | ||
class="math inline">\(P_{LtetO}\)</span> and is repressed by tetR protein. The lower | class="math inline">\(P_{LtetO}\)</span> and is repressed by tetR protein. The lower | ||
| Line 349: | Line 411: | ||
expression levels of the repressor proteins at the beginning is not high enough. | expression levels of the repressor proteins at the beginning is not high enough. | ||
| − | + | </div> | |
| + | </div> | ||
| + | <div id = "cl2" class="demo-card-wide mdl-card mdl-shadow--2dp"> | ||
| + | |||
| + | <div class="mdl-card__supporting-text"> | ||
| + | |||
| + | <h2 id="equations-and-parameters">Equations and Parameters</h2> | ||
<h3 id="repressilator">Repressilator</h3> | <h3 id="repressilator">Repressilator</h3> | ||
The following table lists the reactions in the repressilator network. X, Y, Z represent LacI, | The following table lists the reactions in the repressilator network. X, Y, Z represent LacI, | ||
TetR and cI mRNAs, and PX, PY, PZ represent the corresponding proteins.<br><br> | TetR and cI mRNAs, and PX, PY, PZ represent the corresponding proteins.<br><br> | ||
| − | <div align="center"><img src="https://static.igem.org/mediawiki/2017/9/95/Peking_modelling_clock_table1.png" height="500px"/> | + | <div align="center"><img |
| − | + | src="https://static.igem.org/mediawiki/2017/9/95/Peking_modelling_clock_table1.png" | |
| − | + | height="500px"/> | |
| − | + | </div> | |
| − | + | <br><br> | |
| Line 377: | Line 445: | ||
<h3>Transcription in bio-flip-flop</h3> The transcription kinetics in the bio-flip-flop were | <h3>Transcription in bio-flip-flop</h3> The transcription kinetics in the bio-flip-flop were | ||
| − | + | modified to be repressed by repressors. These can be described by the following | |
| − | + | equations: | |
<br><br> | <br><br> | ||
| − | <div align="center"><img src="https://static.igem.org/mediawiki/2017/9/95/Peking_modelling_clock_table2.png" height="100px"/> | + | <div align="center"><img |
| − | </div><br><br> | + | src="https://static.igem.org/mediawiki/2017/9/95/Peking_modelling_clock_table2.png" |
| + | height="100px"/> | ||
| + | </div> | ||
| + | <br><br> | ||
| + | </div> | ||
| + | </div> | ||
| + | <div id = "cl3" class="demo-card-wide mdl-card mdl-shadow--2dp"> | ||
| + | |||
| + | <div class="mdl-card__supporting-text"> | ||
<h2 id="simulation-results">Simulation Results</h2> | <h2 id="simulation-results">Simulation Results</h2> | ||
We conducted our simulation using Tellurium[^Footnote3], a Python-based simulation platform. | We conducted our simulation using Tellurium[^Footnote3], a Python-based simulation platform. | ||
| − | + | We first adjusted the parameters protein half life and mRNA half life (equivalently | |
| − | + | transcription and translation of repressor proteins) to adjust oscillation period as the one | |
| − | + | observed in the experiment. (此处链接到clock实验)<br><br> | |
| − | <div align="center"><img src="https://static.igem.org/mediawiki/2017/6/66/Peking_modeling_repressilator.png" height="400px"/> | + | <div align="center"><img |
| + | src="https://static.igem.org/mediawiki/2017/6/66/Peking_modeling_repressilator.png" | ||
| + | height="400px"/> | ||
<br><strong>Fig.1. Repressilator simulation results of tetR and cI protein</strong></div> | <br><strong>Fig.1. Repressilator simulation results of tetR and cI protein</strong></div> | ||
<br><br> | <br><br> | ||
| − | + | Then we incorporate the bio-flip-flop into the model, and we got the following simulation | |
| − | + | results: <br><br> | |
| − | <div align="center"><img src="https://static.igem.org/mediawiki/2017/7/73/Peking_modeling_whole_rep_ff.png" height="400px"/> | + | <div align="center"><img |
| + | src="https://static.igem.org/mediawiki/2017/7/73/Peking_modeling_whole_rep_ff.png" | ||
| + | height="400px"/> | ||
<br><strong>Fig.2. Repressilator-driven | <br><strong>Fig.2. Repressilator-driven | ||
| − | frequency divider simulation with two trigger signal groups</strong><br>X_F and Y_F have the same | + | frequency divider simulation with two trigger signal groups</strong><br>X_F and Y_F have |
| − | meaning as those described in Recombinase section. (此处有超链接到第一个model子页面)</div> | + | the same |
| + | meaning as those described in Recombinase section. (此处有超链接到第一个model子页面) | ||
| + | </div> | ||
<br><br> | <br><br> | ||
| − | + | In this simulation | |
| − | + | experiment, the flip-flop lost approximately 10% of its initial state after two trigger | |
| − | + | signal groups. | |
| − | + | ||
| − | + | ||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
</div> | </div> | ||
</div> | </div> | ||
| − | |||
</section> | </section> | ||
| − | + | <section id="Supplementary" class="mdl-components__page mdl-grid"> | |
| − | + | <div class="demo-card-wide mdl-card mdl-shadow--2dp"> | |
| − | + | <div class="mdl-card__supporting-text"> | |
| − | + | <h1>Overview</h1> | |
| − | + | <br> | |
| − | + | <h2 id="pc0">Background</h2> | |
| − | + | In our design, after having designed the flip flop, the device can remember the information of | |
| − | + | its state, the next step is to transform the state into an actual function. To achieve this | |
| − | + | transformation, we first needed a “reader” to read out the current state. At this point the | |
| − | + | control unit comes into play. A control unit is a DNA sequence with recombinase sites whose | |
| − | + | expression is controlled by recom-binase and RDF by reversing or deleting a promoter and/or a | |
| − | + | terminator. To make the control unit reliable and predictable, we first need to be able to | |
| − | + | predict the behaviors of its “building blocks” (or “elements” in electrical engineering), from | |
| − | + | which we weave our engineer’s perspective into the biolog-ical system. However, we need to make | |
| − | + | some adaptations and adjustments to these “elements” to make them usable. | |
| − | + | <br><br> | |
| − | + | </div> | |
| − | + | <!--<div class="mdl-card__actions mdl-card--border"> | |
| − | + | <a class="mdl-button mdl-button--colored mdl-js-button mdl-js-ripple-effect"> | |
| − | + | Get Started | |
| − | + | </a> | |
| − | + | </div>--> | |
| − | + | <div class="mdl-card__menu"> | |
| − | + | <button class="mdl-button mdl-button--icon mdl-js-button mdl-js-ripple-effect"> | |
| − | + | <i class="material-icons">star</i> | |
| − | + | </button> | |
| − | + | </div> | |
| − | + | </div> | |
| − | + | </section> | |
| − | + | </div> | |
</div> | </div> | ||
<section class="docs-text-styling download mdl-color--grey-800" style="height: 1px"></section> | <section class="docs-text-styling download mdl-color--grey-800" style="height: 1px"></section> | ||
Revision as of 06:03, 1 November 2017
Overview
Background
In our design, after having designed the flip flop, the device can remember the information of its state, the next step is to transform the state into an actual function. To achieve this transformation, we first needed a “reader” to read out the current state. At this point the control unit comes into play. A control unit is a DNA sequence with recombinase sites whose expression is controlled by recom-binase and RDF by reversing or deleting a promoter and/or a terminator. To make the control unit reliable and predictable, we first need to be able to predict the behaviors of its “building blocks” (or “elements” in electrical engineering), from which we weave our engineer’s perspective into the biolog-ical system. However, we need to make some adaptations and adjustments to these “elements” to make them usable.Control Unit
Intention of the Model
In this model, we aim to describe the sequence modification process of the control unit sequence with recombination sites. Different recombinases are expressed in series to alter the direction of terminators between corresponding recombination sites on the control unit circuit. The combination of unidirectional terminators in different directions (reverse or foward) determines the state of RNA polymerase flux, thus controlling expression of the downstream gene.Though we have done simulation of recombinase dynamics in the previous section, it is necessary to consider two key concepts of biological processes:
1. population behavior 2. stochastic process
The reason for modeling cell population behavior is that deployment of synthetic bacterial devices would certainly involve populations of cells. Furthermore, we demonstrate our design through continuous imaging of entire cell cultures. This is a theory-experiment interaction procedure. The reason for stochastic modeling is self-evident. Cellular biological processes, especially gene expression, are innately noisy and stochastic.[^Footnote1] Previous work has demonstrated the effectiveness of the integration of population-level dynamics and stochastic cellular responses to inputs.[^Footnote2]It is important to explain how stochastic single-cell responses affect overall population-level distributions and behavior.
This section describes a Markov model of recombinase-based DNA flipping and a Gillespie stochastic simulation method[^Footnote3] to observe theoretical sequence state evolution. We will focus on our demonstration design, which can be generalized to other designs.
Biological Basis
The control units are implemented experimentally by transforming a plasmid into an E. coli. This plasmid has similar structure as the reporter plasmid in the previous section. However, in this situation the sequence fragments being inverted are terminators. Using experimentally screened uni-directional terminators, the transcription or polymerase flux[^Footnote4] can be precisely controlled. The figure below illustrates the result of the design software. (超链接到software)

Clock
Intention of the Model
We attempts to develop a framework of biological sequential circuits that are programmable. The envisioned circuit is capable of both storing states in DNA and automatically running a series of instructions in a specific order. According to our design, our bio-flip-flop has two promoters, each triggered by an inducing signal. Therefore, if we would like to build an automatic triggering system for the bio-flip-flop, we need at least two signals in our clock. It is then self-evident to incorporate the classic structure in synthetic biology: the repressilator.This model serves as a proof of concept for such implementation. We first constructed an ordinary differential equation (ODE) system to describe the repressilator, then we used two repressor proteins expressed by the repressilator to regulate the two promoters. Finally we adjusted parameters and gave a simulation result as a proof of the possibility of repressilator-driven bio-flip-flop.
Biological Basis and Assumptions
The repressilator is a synthetic genetic regulatory network consisting of a ring-oscillator with three genes, each expressing a protein that represses the next gene in the loop.[^Footnote1] This network was designed from scratch to exhibit a stable oscillation which is reported via the expression of green fluorescent protein, and hence acts like an electrical oscillator system with fixed time periods. The repressilator consists of three genes connected in a feedback loop, such that each gene represses the next gene in the loop, and is repressed by the previous gene.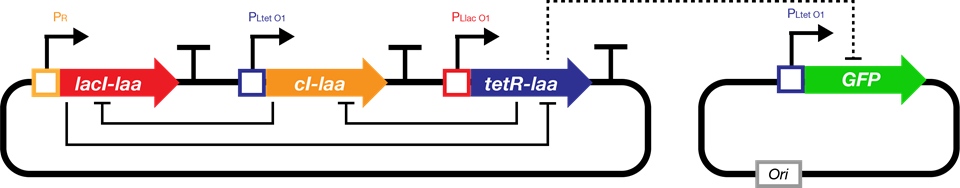
Fig.1. The genetic structure of the repressilator network
We decided to implement our repressilator-triggered bio-flip-flop with two repressor proteins regulating the two promoters’ transcription. The basis of ODE system construction is illustrated as follows:
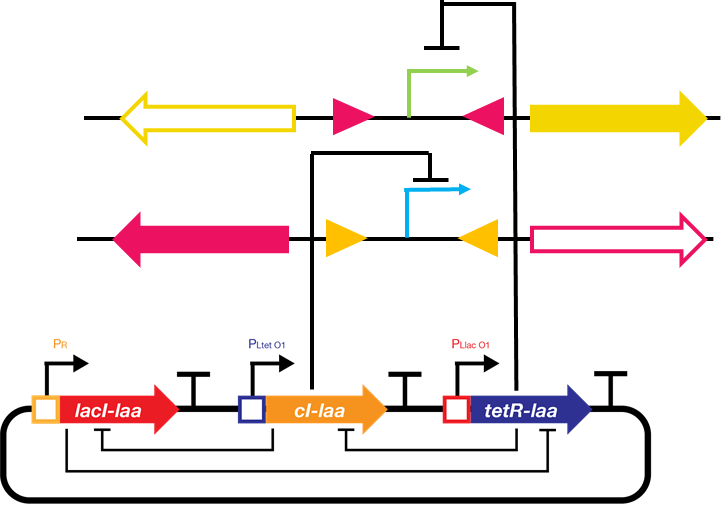
Fig.2. Schematic drawing of the repressilator-driven bio-flip-flop
The upper promoter (X) is \(P_{LtetO}\) and is repressed by tetR protein. The lower promoter (Y) is \(P_{R}\) and is repressed by cI protein.Simulation of bio-flip-flop starts at a later time than repressilator because the expression levels of the repressor proteins at the beginning is not high enough.
Equations and Parameters
Repressilator
The following table lists the reactions in the repressilator network. X, Y, Z represent LacI, TetR and cI mRNAs, and PX, PY, PZ represent the corresponding proteins.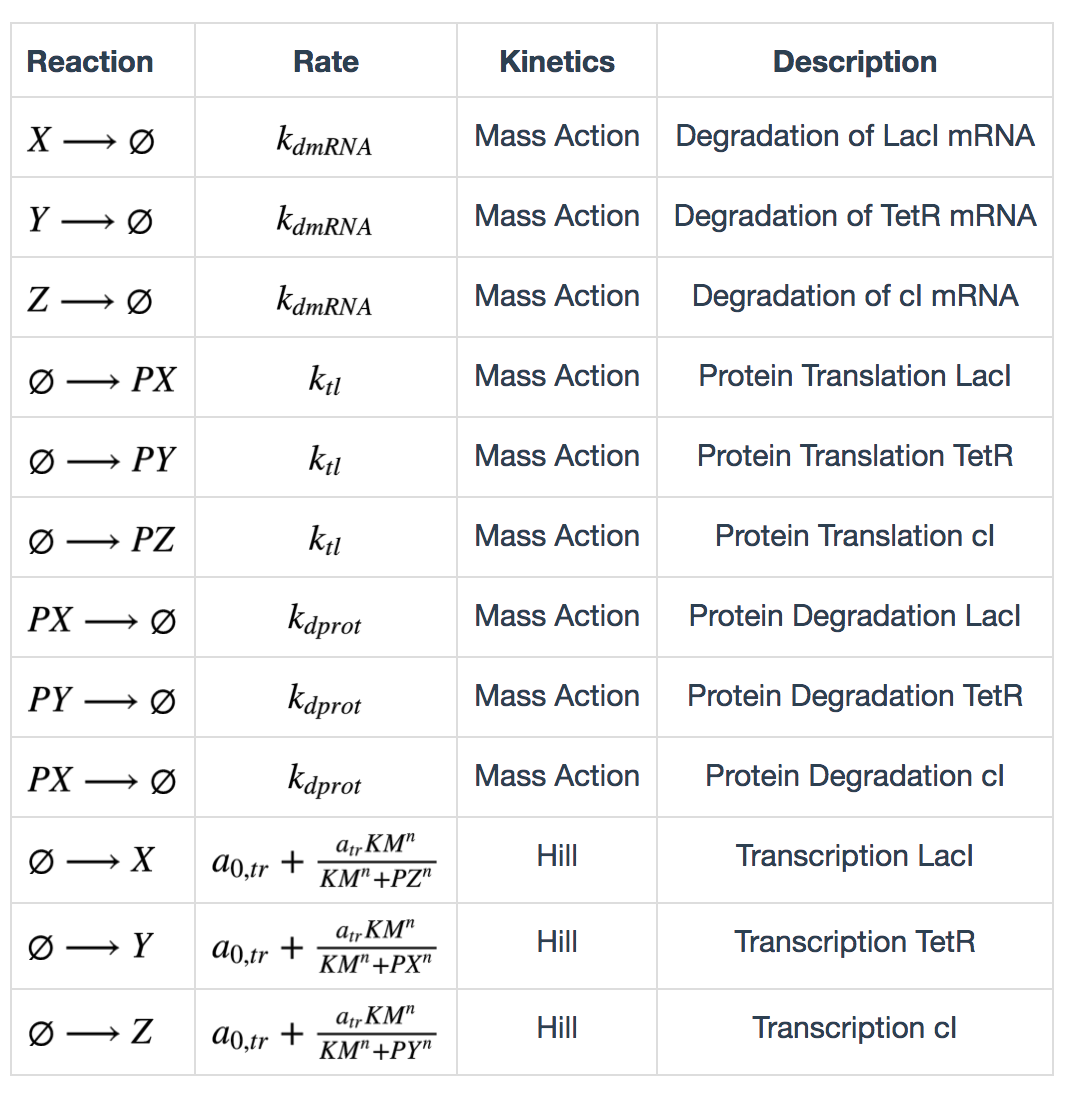
Calculation of degradation rates are described as follows: \[ k_{dmRNA} = \frac{ln(2)}{\tau_{mRNA}} \] \[ k_{dprot} = \frac{ln(2)}{\tau_{protein}} \] \(\tau_{mRNA}\) and \(\tau_{protein}\) are half lives of mRNA and protein. Calculation of maximum transcription rate: \[ a_{tr} = 60(ps_a - ps_0) \] \(ps_a\) is “tps active” and \(ps_0\) is “tps_repressive”. Calculation of translation rate: \[ k_{tl} = \frac{eff}{t_{ave}} \] \(eff\) is translation efficiency and \(t_{ave}\) is average mRNA life time. \(t_{ave} = \frac{\tau_{mRNA}}{ln(2)}\)
Transcription in bio-flip-flop
The transcription kinetics in the bio-flip-flop were modified to be repressed by repressors. These can be described by the following equations: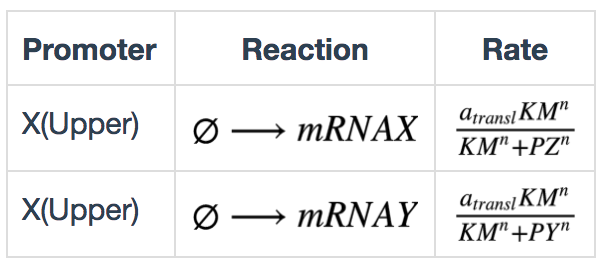
Simulation Results
We conducted our simulation using Tellurium[^Footnote3], a Python-based simulation platform. We first adjusted the parameters protein half life and mRNA half life (equivalently transcription and translation of repressor proteins) to adjust oscillation period as the one observed in the experiment. (此处链接到clock实验)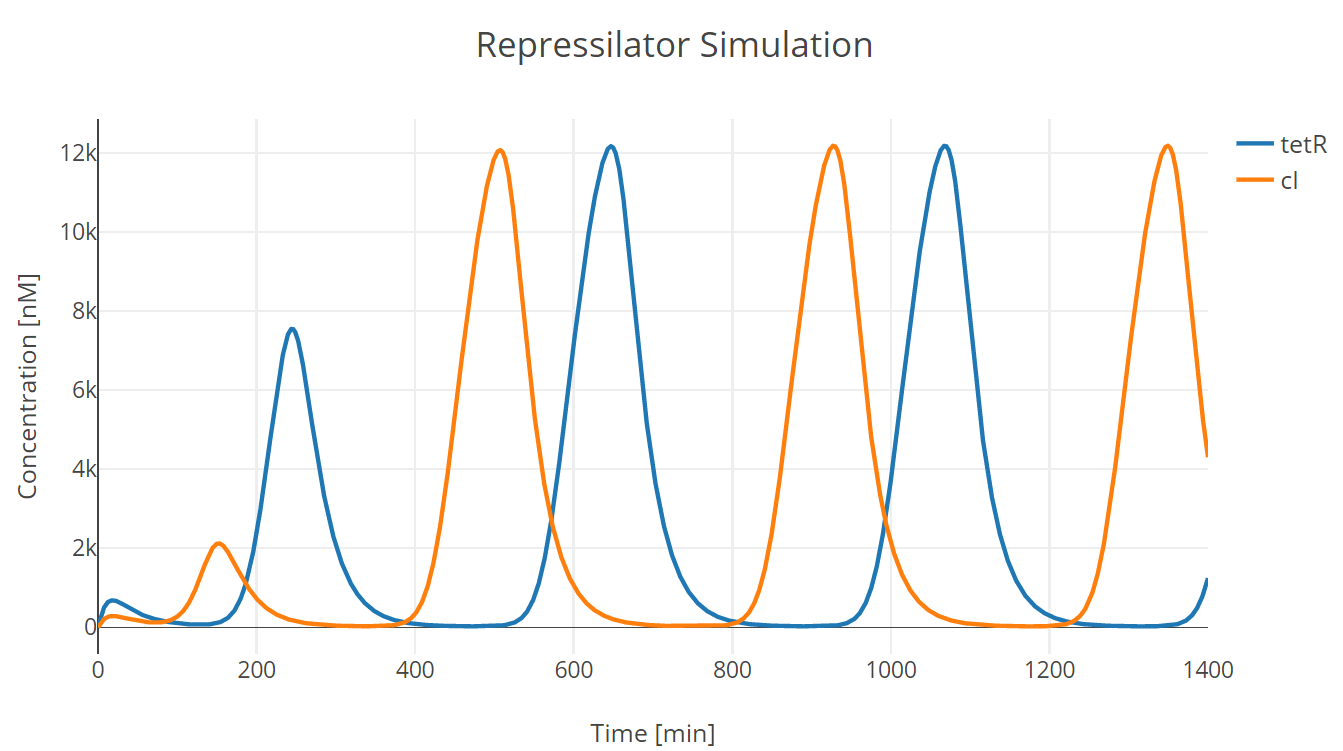
Fig.1. Repressilator simulation results of tetR and cI protein
Then we incorporate the bio-flip-flop into the model, and we got the following simulation results:
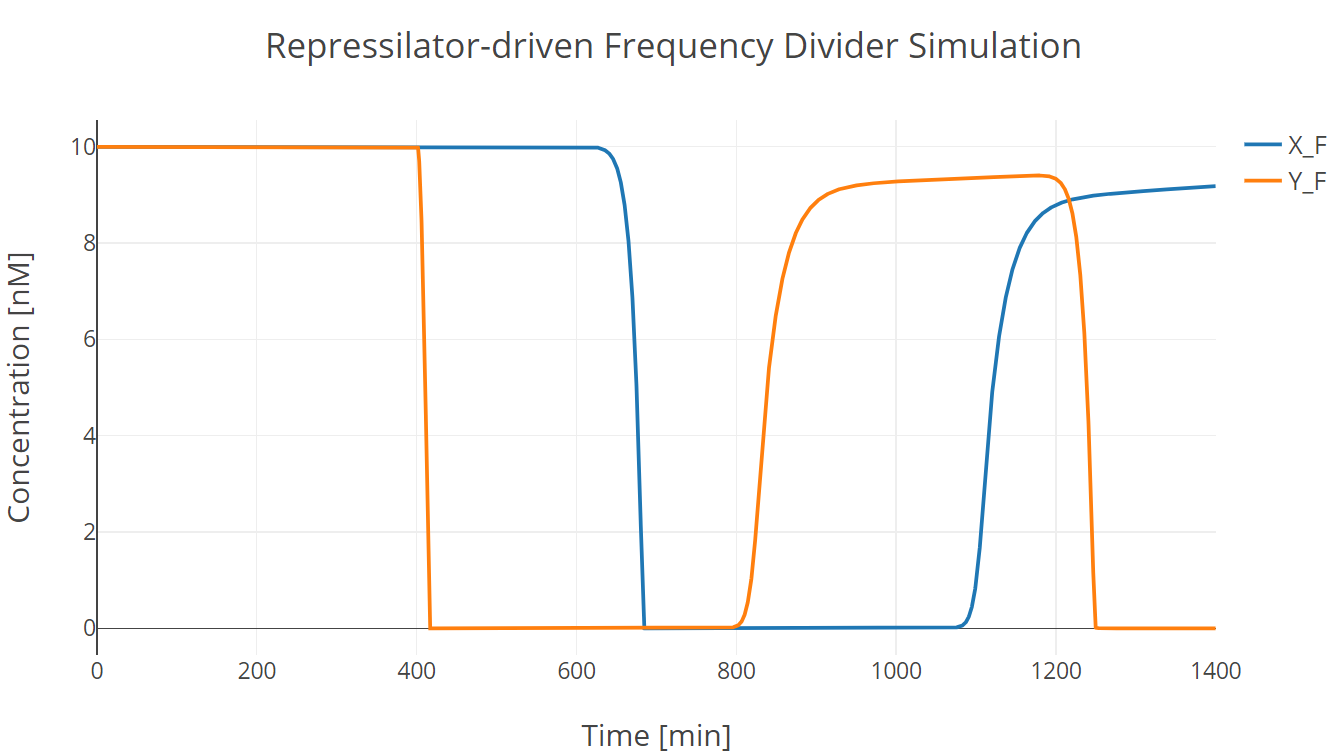
Fig.2. Repressilator-driven frequency divider simulation with two trigger signal groups
X_F and Y_F have the same meaning as those described in Recombinase section. (此处有超链接到第一个model子页面)
In this simulation experiment, the flip-flop lost approximately 10% of its initial state after two trigger signal groups.