Team:Heidelberg/Model/Medium Consumption
Medium consumption model
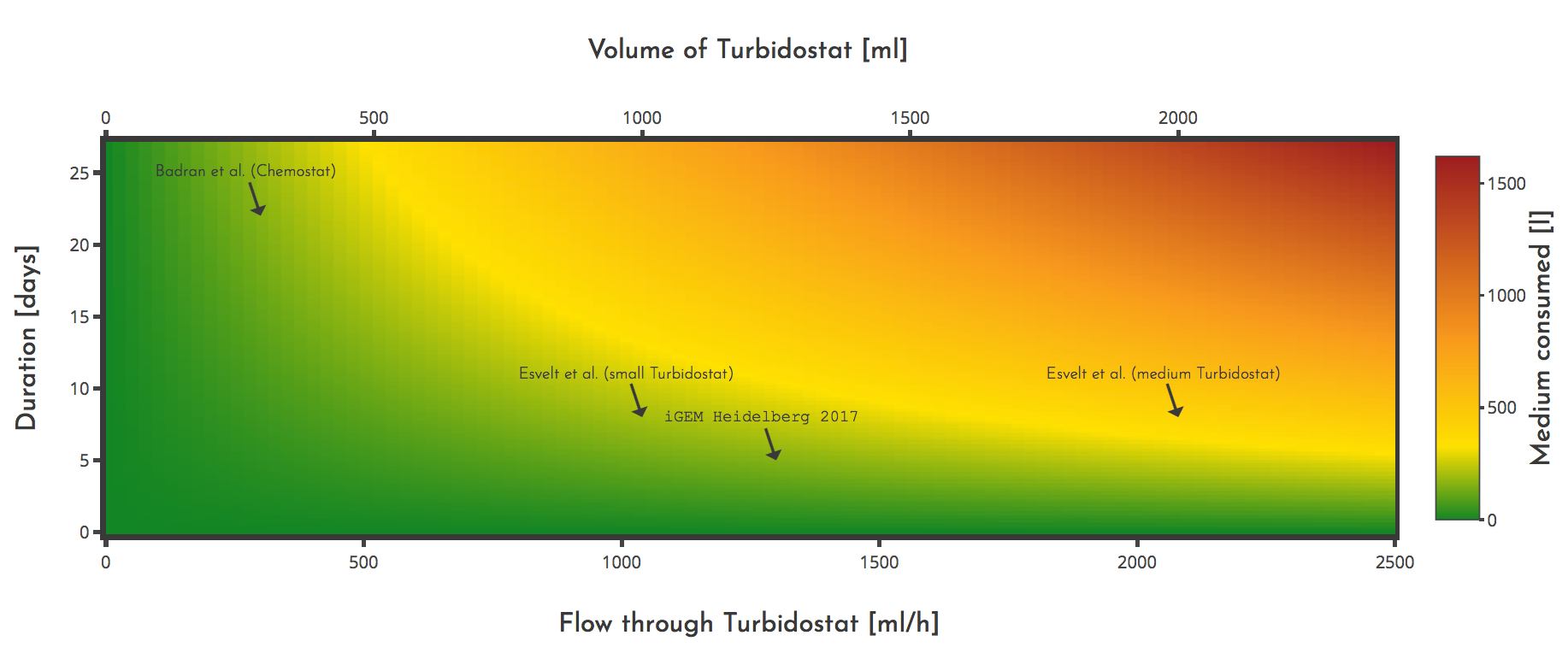
PACE usually consumes an extraordinary amount of medium per experiment. This is due to the need for a continuous supply of host cells with a constant cell density. This can be achieved either by using a turbidostat or a chemostat. Here we provide a tool to both the community and ourselves to calculate medium consumption based on different tunable parameters of PACE. We also want to gain an understanding of how we can reduce the amount of medium needed for an experiment.Medium consumption is critical when it comes to the energy needed for an experiment especially because the medium has to be autoclaved, which is highly energy consuming. In a turbidostat, the cell density is held constant by adjusting the medium influx to the cell density. That means the growth of the cells is not affected, instead for a one new cell, emerging from proliferation, one is put to the waste. In a Chemostat, the cell density is controlled by adjusting the influx of an essential nutrient, which limits the growth of the culture to a certain cell density. This may cause the host cells to be less efficient in producing the proteins needed for PACE and replicating the phage genome. Additionally, there is a constant efflux from the chemostat to the lagoons, that compensates the growth of the cells.
iGEM Heidelberg used a turbidostat in all PACE experiments because we assumed that PACE works better when the E. coli cells are allowed to grow at their maximum rate under the given conditions. So their ability to produce phage as fast as possible is not impaired by any deprivation.
The flow rate through a turbidostat is described by $$ \frac{\partial V_{M} }{\partial t} = \Phi_{T} = \frac{log(2)}{t_{E} } \cdot V_{T} $$ The flow rate \(\Phi_{T}\) in has to be equal to the growth term normalized with the turbidostat size. Note that the total volume of the turbidostat is constant because the flow rate is the same for incoming and outgoing medium.
Table 1: Additional Variables and Parameters used in the numeric solution of the model. List of all paramters and variables used in the analytic solution of this model and in the corresponding interactive webtool
| Symbol | Value and Unit | Explanation |
|---|---|---|
| \(V_{T}\) | [ml] | Volume of Turbidostat |
| \(V_{M}\) | [ml] | Volume of Medium consumed |
| \(t_{E} \) | [min] | E. coli generation time |
| \(\Phi_{T}\) | [ml/h] | Flow rate through turbidostat |
| \(t_{max}\) | [min] | Duration of the experiment |
Minmal Turbidostat Volume
Larger turbidostats or chemostats with a larger flow needs more medium for the same duration than smaller ones. When working with the minimal required volume or flow you can save medium and thus also energy. The minimal flow that is required can be calculated using $$ \Phi_{T} = b \cdot V_{L} \cdot N_{L} \cdot \Phi_{L} $$ which is simply the product of the lagoon volume, count and flow rate and a safety factor \(b\).In case of fluctuations in the generation time of the E. coli cells it is crucial to have a buffer so that the turbidostat is not diluted when the culture grows slower. A buffer of 50 %, means \(b\) is set to \(1.5\). For a turbidostat, the volume can be calculated from the flow using $$ V_{T} = \Phi_{T} \cdot \frac{t_{E} }{log(2)} $$ For chemostats only the needed flow rate \(\Phi_{T}\) is calculated, the volume needs to be chosen so that the flow rate can be reached.
Table 2: Additional Variables and Parameters used for this calculation. List of all additional paramters and variables used in the analytic solution of this model and in the corresponding interactive webtool
| Symbol | Value and Unit | Explanation |
|---|---|---|
| \(V_{L}\) | [ml] | Volume of Lagoons |
| \(N_{L}\) | Number of Lagoons | |
| \(\Phi_{L}\) | [ml/min] | E. coli generation time |
| \(b\) | \(1.5\) | Buffer |
Minimal Lagoon Volume
Obviously smaller lagoons require smaller turbidostats or chemostats with a lower flow rate and are therefore saving medium. However, there is a lower limit to lagoon size, if the phage population is too small, the sequence space that can be covered is insufficient to find variants that are better than previous ones. Lagoon sizes used by other vary from 15 mlTable 3: Additional Variables and Parameters used for this calculation. List of all additional paramters and variables used in the analytic solution of this model and in the corresponding interactive webtool
| Symbol | Value and Unit | Explanation |
|---|---|---|
| \(N_{P}\) | [pfu] | Amount of phages per lagoon |
| \(c_{P}\) | [pfu/ml] | Phage concentration |
| \(L_{S}\) | [bp] | Sequence length in basepairs |
| \(L_{T}\) | [bp] | Total sequence length in basepairs in lagoon |
| \(N_{M}\) | Number of mutations | |
| \(r_{M}\) | [1/generation] | Number of mutated basepairs per basepair per generation |
| \(n\) | [bp] | Number of mutated basepairs |
| \(M_{n}\) | Number of real sequences with \(n\) mutations | |
| \(N_{n}\) | Number of possible sequences with \(n\) mutations | |
| \(t\) | Theortical coverage of double |
Practice
The PACE experiments we carried out ranged from 3 to 7 days with a turbidostat volume of about 1.2 l. We observed E. coli generation times of 30 to 40 min.